Do snakes have a third eye? - Reptilenesia
 |
| Limerick written by Annie Simminger about her nephew, Richard Marshall Eakin, and published in his 1983 book The Third Eye. Eakin and Robert C. Stebbins performed and published many experiments on the structure and function of the third eyes of lizards |
Well, they're not nostrils. I don't think this has been scientifically evaluated before but maybe sensory related. Let's ask @SssnakeySci. https://t.co/uwfvdLuYwI— David Steen, Ph.D. (@AlongsideWild) January 17, 2017
 |
| Parietal eye (black outline) and parietal scale (white outline) of Liolaemus bisignatus (Philippi's Tree Iguana). From Labra et al. 2010 |
 |
| The "chimney-like" pineal foramen of the extinct 3-foot-long 250 million year old South African fossil therapsid Hipposaurus. From Haughton 1929 |
 |
| Endocast of the brain of the dog-sized 250-million-year-old dicynodont Lystrosaurus, the "humble badass of the Triassic", showing the large parietal eye (dark structure at the top). From Edinger 1955 |
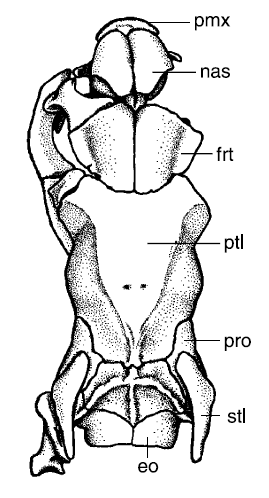 |
| Paired parietal foramina in the parietal (ptl) bone of a Banded Krait (Bungarus fasciatus) skull. From Scanlon & Lee 2004 |
 |
| Developmental origins of the parietal ("median") eye and the lateral eye. The cilia are cellular structures that normally function for movement (e.g., of debris out of the nose, of water over gills, of eggs into oviducts, of sperm cells to the egg). In the eye, they have evolved into photoreceptors. |
 |
| Diagram of the lizard parietal eye From Solessio & Engbretson 1993 |
 |
| Comparative morphology of the pineal complex in A) lamprey, B) frog, C) lizard, and D) human. From Edinger 1955 |
 |
| The parietal eye of a Western Fence Lizard (Sceloporus occidentalis) C = cornea; CC = connective tissue; L = lumen; LS = lens; PN = parietal nerve; R = retina Light micrograph from Eakin 1970 |
 |
| Parietal spots of a Copperhead (Agkistrodon contortrix) |


1 The function of vitamin A in eyesight was the basis for a WWII propaganda campaign that eating more carrots could improve human night vision. Although it's true that carrots and vitamin A are essential for good eyesight, the extent to which eating more carrots can improve a person's eyesight was apparently greatly exaggerated in 1940 to create a cover story for the novel abilities of Allied pilots to pinpoint Axis fighter jets at night, which in reality was due to on-board Airborne Interception Radar (although there is in turn some disagreement among historians as to how purposeful the deception was and how much both sides knew about the other side's radar capabilities).↩
2 Melatonin is synthesized from the amino acid tryptophan, which is the origin of another common myth: that eating a ton of turkey causes you feel sleepy.↩
3 Tilly Edinger was among the very last scientists of Jewish ancestry to leave pre-WWII Germany. A 1938 letter to the U. S. State Department in support of her immigration application from George Gaylord Simpson read "She is a research scientist of the first rank and is favorably known as such all over the world. She is everywhere recognized as the leading specialist on the study of the brain and nervous system of extinct animals and on the evolution of the gross structure of the brain. She is so preeminent in this field that she may really be said to have created a new branch of science, that of paleo-neurology, a study of outstanding value and importance”. She was the first female president of the Society of Vertebrate Paleontology, and authored over 1200 scientific papers and books, many sprinkled with sharp-witted, humorous phrases and observations. Her pioneering work in paleoneurology is well-chronicled here.↩
4 During embryonic development, the parietal eye and the pineal organ form together from a pocket formed in the brain ectoderm. The ancestral state is presumed to have been a possibly paired photosensory organ, as seen in extant lampreys. The parietal eye and the pineal gland of tetrapods are probably the descendants of the left and right parts of this organ, respectively. Some Devonian fishes have two parietal foramina in their skulls, suggesting an ancestral bilaterality of parietal eyes.↩
5 Crocodilians and some tropical lineages of mammals (some xenarthrans [sloths], pangolins, sirenians [manatees & dugongs], some marsupials [sugar gliders]) have lost both their parietal eye and their pineal organ. All amphibians have a pineal organ, but some frogs and toads also have what is called a "frontal organ", which is essentially a parietal eye. The word "pineal" comes from the shape of the human pineal organ, which resembles a pine cone.↩
Adler, K. and J. B. Phillips. 1985. Orientation in a desert lizard (Uma notata): time-compensated compass movement and polarotaxis. Journal of Comparative Physiology A 156:547-552 <link>
Arendt, J. 1994. Melatonin and the Mammalian Pineal Gland. Springer Science & Business Media <link>
Benoit, J., P. Manger, and B. Rubidge. 2016. Palaeoneurological clues to the evolution of defining mammalian soft tissue traits. Scientific Reports 6 <link>
Bertolucci, C., A. Foa, and G. Tosini. 2002. The circadian organization of reptiles. Pages 129-143 in V. Kumar, editor. Biological Rhythms. Springer <preview>
Bradshaw, W. E. and C. M. Holzapfel. 2007. Evolution of animal photoperiodism. Annual Review of Ecology, Evolution, and Systematics 38:1-25 <link>
Braun-Elwert, C. 2011. Tuatara and their living fossil label. Master of Science Communication thesis. University of Otago, Dunedin, New Zealand <link>
Buchholtz, E. A. and E.-A. Seyfarth. 1999. The gospel of the fossil brain: Tilly Edinger and the science of paleoneurology. Brain Research Bulletin 48:351-361 <link>
Caprette, C. L., M. S. Y. Lee, R. Shine, A. Mokany, and J. F. Downhower. 2004. The origin of snakes (Serpentes) as seen through eye anatomy. Biological Journal of the Linnean Society 81:469-482 <link>
Crews, D., V. Hingorani, and R. J. Nelson. 1988. Role of the pineal gland in the control of annual reproductive behavioral and physiological cycles in the red-sided garter snake (Thamnophis sirtalis parietalis). Journal of Biological Rhythms 3:293-302 <abstract>
Dendy, A. 1911. On the structure, development and morphological interpretation of the pineal organs and adjacent parts of the brain in the tuatara (Sphenodon punctatus). Philosophical Transactions of the Royal Society of London. Series B, Containing Papers of a Biological Character 201:227-331.
Dodt, E. 1973. The parietal eye (pineal and parietal organs) of lower vertebrates. Pages 113-140 Visual Centers in the Brain. Springer.
Eakin, R. M. 1970. A Third Eye: A century-old zoological enigma yields its secrets to electron-microscopist and neurophysiologist. American Scientist 58:73-79 <abstract>
Eakin, R. M. 1973. The Third Eye. University of California Press <Google Book>
Eakin, R. M. and J. A. Westfall. 1960. Further observations on the fine structure of the parietal eye of lizards. The Journal of Cell Biology 8:483-499.
Edinger, T. 1955. The size of parietal foramen and organ in reptiles: a rectification. Bulletin of the Museum of Comparative Zoology 114:1-34 <link>
Engbretson, G. A. 1992. Neurobiology of the lacertilian parietal eye system. Ethology Ecology & Evolution 4:89-107 <abstract>
Foster, R., J. Garcia-Fernandez, I. Provencio, and W. DeGrip. 1993. Opsin localization and chromophore retinoids identified within the basal brain of the lizard Anolis carolinensis. Journal of Comparative Physiology A 172:33-45.
Foster, R. G., M. S. Grace, I. Provencio, W. J. Degrip, and J. Garcia-Fernandez. 1994. Identification of vertebrate deep brain photoreceptors. Neuroscience & Biobehavioral Reviews 18:541-546.
Foureaux, G., M. I. Egami, C. Jared, M. M. Antoniazzi, R. C. Gutierre, and R. L. Smith. 2010. Rudimentary eyes of squamate fossorial reptiles (Amphisbaenia and Serpentes). The Anatomical Record 293:351-357 <link>
Freake, M. J. 2001. Homing behaviour in the sleepy lizard (Tiliqua rugosa): the role of visual cues and the parietal eye. Behavioral Ecology and Sociobiology 50:563-569 <link>
Gundy, G. C. and G. Z. Wurst. 1976. Parietal eye‐pineal morphology in lizards and its physiological implications. The Anatomical Record 185:419-431 <abstract>
Gundy, G. C. and G. Z. Wurst. 1976. The occurrence of parietal eyes in recent Lacertilia (Reptilia). Journal of Herpetology 10:113-121 <abstract>
Haldar, C. and R. Pandey. 1989. Effect of pinealectomy on annual testicular cycle of Indian chequered water snake, Natrix piscator. General and Comparative Endocrinology 76:214-222 <abstract>
Haldar, C. and R. Pandey. 1989. Effect of pinealectomy on the testicular response of the freshwater snake Natrix piscator to different environmental factors. Canadian Journal of Zoology 67:2352-2357 <link>
Hamasaki, D. and D. Eder. 1977. Adaptive radiation of the pineal system. Pages 497-548 The Visual System in Vertebrates. Springer <Google Book>
Haughton, S. 1929. On some new therapsid genera. Annals of the South African Museum 28:55-78 <link>
Hawley, A. W. and M. Aleksiuk. 1976. The influence of photoperiod and temperature on seasonal testicular recrudescence in the red-sided garter snake (Thamnophis sirtalis parietalis). Comparative Biochemistry and Physiology Part A: Physiology 53:215-221.
Hsiang, A. Y., D. J. Field, T. H. Webster, A. D. Behlke, M. B. Davis, R. A. Racicot, and J. A. Gauthier. 2015. The origin of snakes: revealing the ecology, behavior, and evolutionary history of early snakes using genomics, phenomics, and the fossil record. BMC Evolutionary Biology 15:1-22 <link>
Ireland, L. C. and C. Gans. 1977. Optokinetic behavior of the tuatara, Sphenodon punctatus. Herpetologica 33:339-344.
Jenison, G. and J. Nolte. 1980. An ultraviolet-sensitive mechanism in the reptilian parietal eye. Brain Research 194:506-510
Kitay, J. I. and M. D. Altschule. 1954. The pineal gland. A review of the physiologic literature. Harvard Press, Cambridge, Massachusetts.
Labra, A., K. L. Voje, H. Seligmann, and T. F. Hansen. 2010. Evolution of the third eye: a phylogenetic comparative study of parietal‐eye size as an ecophysiological adaptation in Liolaemus lizards. Biological Journal of the Linnean Society 101:870-883 <link>
Leydig, F. 1872. Zur Kenntniss der Sinnesorgane der Schlangen. Archiv für mikroskopische Anatomie 8:317-357 <abstract>
Linder, H. 1913. Beiträge zur Kenntnis der Plesiosaurier-Gattungen Peloneustes und Pliosaurus. Neues Jahrbuch für Geologie und Paläontologie–Abhandlungen 15:337-409.
Lutterschmidt, D. I., W. I. Lutterschmidt, and V. H. Hutchison. 1997. Melatonin and chlorpromazine: thermal selection and metabolic rate in the bullsnake, Pituophis melanoleucus. Comparative Biochemistry and Physiology Part C: Pharmacology, Toxicology and Endocrinology 118:271-277 <abstract>
Lutterschmidt, D. I., W. I. Lutterschmidt, and V. H. Hutchison. 2003. Melatonin and thermoregulation in ectothermic vertebrates: a review. Canadian Journal of Zoology 81:1-13 <link>
Lutterschmidt, D. I., W. I. Lutterschmidt, N. B. Ford, and V. H. Hutchison. 2002. Behavioral thermoregulation and the role of melatonin in a nocturnal snake. Hormones and Behavior 41:41-50 <link>
Menaker, M. and G. Tosini. 1996. The evolution of vertebrate circadian systems. Pages 39-52 in K. Honma and S. Honma, editors. Sapporo Symposium on Biological Rhythms: Circadian Organization and Oscillatory Coupling. Hokkaido University Press, Sapporo, Japan.
Menaker, M. and S. Wisner. 1983. Temperature-compensated circadian clock in the pineal of Anolis. Proceedings of the National Academy of Sciences 80:6119-6121 <link>
Mendonça, M. T., A. J. Tousignant, and D. Crews. 1995. Seasonal changes and annual variability in daily plasma melatonin in the red-sided garter snake (Thamnophis sirtalis parietalis). General and Comparative Endocrinology 100:226-237 <link>
Mendonca, M. T., A. J. Tousignant, and D. Crews. 1996. Pinealectomy, melatonin, and courtship behavior in male red‐sided garter snakes (Thamnophis sirtalis parietalis). The Journal of Experimental Zoology 274:63-74 <link>
Mendonça, M. T., A. J. Tousignant, and D. Crews. 1996. Courting and noncourting male red-sided garter snakes, Thamnophis sirtalis parietalis: Plasma melatonin levels and the effects of pinealectomy. Hormones and Behavior 30:176-185 <link>
Miller, W. H. and M. L. Wolbarsht. 1962. Neural activity in the parietal eye of a lizard. Science 135:316-317 <abstract>
Moore, A. F. and M. Menaker. 2011. The effect of light on melatonin secretion in the cultured pineal glands of Anolis lizards. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 160:301-308 <abstract>
Nelson, R. J., R. T. Mason, R. W. Krohmer, and D. Crews. 1987. Pinealectomy blocks vernal courtship behavior in red-sided garter snakes. Physiology & Behavior 39:231-233 <link>
Petit, A. 1968. Embryogénèse de l'épiphyse et de l'organe sous-commissural de la couleuvre à collier (Tropidonotus natrix L.). Arch. Anat. (Strasbourg) 52:3-25.
Petit, A. 1971. L'épiphyse d'un serpent: Tropidonotus natrix L. Zeitschrift für Zellforschung und mikroskopische Anatomie 120:94-119 <abstract>
Quay, W. 1979. The parietal eye–pineal complex. Pages 245-406 in C. Gans, R. G. Northcutt, and P. Ulinski, editors. Biology of the Reptilia. Volume 9. Neurology A. Academic Press, London <link>
Quay, W., J. A. Kappers, and J. Jongkind. 1968. Innervation and fluorescence histochemistry of monoamines in the pineal organ of a snake (Natrix natrix). Journal of Neuro-visceral Relations 31:11-25 <abstract>
Ralph, C. 1975. The pineal gland and geographical distribution of animals. International Journal of Biometeorology 19:289-303.
Ralph, C. L., B. T. Firth, and J. S. Turner. 1979. The role of the pineal body in ectotherm thermoregulation. American Zoologist 19:273-293 <link>
Ralph, C. L., S. Young, R. Gettinger, and T. O'Shea. 1985. Does the manatee have a pineal body? Acta Zoologica 66:55-60 <abstract>
Scanlon, J. D. and M. S. Lee. 2004. Phylogeny of Australasian venomous snakes (Colubroidea, Elapidae, Hydrophiinae) based on phenotypic and molecular evidence. Zoologica Scripta 33:335-366 <link>
Seebacher, F. and S. A. Murray. 2007. Transient receptor potential ion channels control thermoregulatory behaviour in reptiles. PLoS ONE 2:e281 <link>
Simões, B. F., F. L. Sampaio, C. Jared, M. M. Antoniazzi, E. R. Loew, J. K. Bowmaker, A. Rodriguez, N. S. Hart, D. M. Hunt, J. C. Partridge, and D. J. Gower. 2015. Visual system evolution and the nature of the ancestral snake. Journal of Evolutionary Biology 28:1309-1320 <link>
Solessio, E. and G. A. Engbretson. 1993. Antagonistic chromatic mechanisms in photoreceptors of the parietal eye of lizards. Nature 364:442-445 <abstract>
Stebbins, R. C. 1958. An experimental study of the" third eye" of the tuatara. Copeia 1958:183-190 <abstract>
Stebbins, R. C. and R. M. Eakin. 1958. The role of the" third eye" in reptilian behavior. American Museum Novitates 1870:1-40 <link>
Tilden, A. R. and V. H. Hutchison. 1993. Influence of photoperiod and temperature on serum melatonin in the diamondback water snake, Nerodia rhombifera. General and Comparative Endocrinology 92:347-354 <abstract>
Tosini, G. 1997. The pineal complex of reptiles: physiological and behavioral roles. Ethology Ecology & Evolution 9:313-333 <link>
Tosini, G., C. Bertolucci, and A. Foà. 2001. The circadian system of reptiles: a multioscillatory and multiphotoreceptive system. Physiology & Behavior 72:461-471 <abstract>
Trost, E. 1952. Untersuchungen über die frühe Entwicklung des Parietalauges und der Epiphyse von Anguis fragilis, Chalcides ocellatus und Tropidonotus natrix. Zoologischer Anzeiger 148:58-71.
Underwood, H. and M. Menaker. 1976. Extraretinal photoreception in lizards. Photochemistry and Photobiology 23:227-243 <abstract>
Underwood, H. 1989. The pineal and melatonin: regulators of circadian function in lower vertebrates. Cellular and Molecular Life Sciences 45:914-922 <link>
Ung, C. Y. J. and A. C. Molteno. 2004. An enigmatic eye: the histology of the tuatara pineal complex. Clinical & Experimental Ophthalmology 32:614-618 <abstract>
Vivien-Roels, B., P. Pévet, M. Dubois, J. Arendt, and G. Brown. 1981. Immunohistochemical evidence for the presence of melatonin in the pineal gland, the retina and the Harderian gland. Cell and Tissue Research 217:105-115.
Vollrath, L. 1979. Comparative morphology of the vertebrate pineal complex. Progress in Brain Research 52:25-38 <Google Book>
Wurst, G. and G. Gundy. 1982. Pineal morphology in amphisbaenians. Page 908 in Annual Meeting of the American Society of Zoologist. American Society of Zoologists, Louisville, Kentucky.
Zimmerman, K. and H. Heatwole. 1990. Cutaneous photoreception: a new sensory mechanism for reptiles. Copeia 1990:860-862.




